Glycobiology of model organisms
The sequencing of the genomes of multicellular organisms has shown that 1% of all genes are directly relevant to glycoconjugate metabolism.
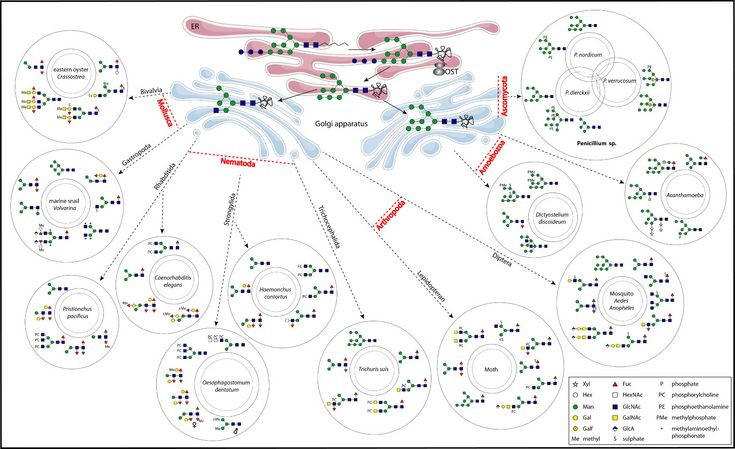
Over the years we have cloned cDNAs encoding a number of glycosyltransferases from Arabidopsis thaliana, rice, Caenorhabditis elegans and Drosophila melanogaster with a particular focus on fucosyltransferases (see, e.g., J. Biol. Chem., 276, 28058-67, J. Biol. Chem., 279, 49588-98, J. Biol. Chem., 281, 3343-53 and J. Biol. Chem. 288, 21015-28). These enzymes catalyse the attachment of fucose to glycans – in particular we have focused on those modifying the N-glycan core region; they can then be used as tools for generating fucosylated glycans in vitro.
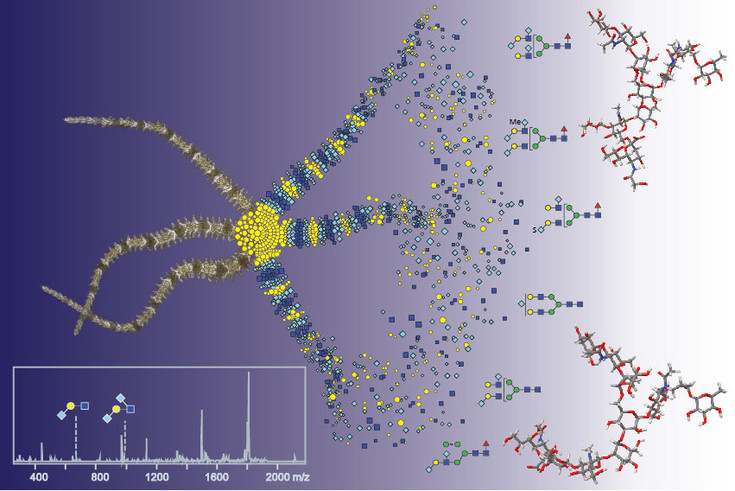
Fucose is a sugar which is often associated with recognition events – such as selectin-mediated interactions during inflammatory responses. In the context of plant glycosylation, core α1,3-fucose is present on many allergens and is immunogenic; in Caenorhabditis and Drosophila the same structure is present in neuronal tissue. Indeed antibodies raised against a plant glycoprotein, horseradish peroxidase (HRP), recognise neural tissue in many invertebrates. In order to define which fucosyltransferases are responsible in vivo for the formation of this anti-HRP epitope, we have performed glycan analysis of mutant Caenorhabditis and used RNAi in a Drosophila neural cell line. A mix of biochemical and genetic approaches is now being used to examine glycosylation in Dictyostelium discoideum (Biochem. J., 423, 41-52 and J. Prot. Res. 12, 1173-87).
Complementary to the study of fucosyltransferases is the examination of the enzymes responsible for making the fucose donor, GDP-Fucose, and other enzymes (such as hexosaminidases) which play roles in glycan biosynthesis. In the mouse, we have uncovered a novel hexosaminidase (HexD; Biochem. J., 419, 83-90) and also examined the closest worm relatives of HexD using a range of techniques (J. Biol. Chem., 282, 27825-40 and J. Biol. Chem., 287, 28276–90). We also examined the Golgi mannosidase II from Caenorhabditis (J. Biol. Chem., 281, 28265-77) which is a key enzyme for normal N-glycan biosynthesis not just in worms but also in mammals.
Most recently, we have investigated a novel GalNAc-specific hexosaminidase from Caenorhabditis (J. Biol. Chem., 299, 103053) and have shown alterations in the N-glycome of the model worm during development or due to cultivation method (Mol. Cell. Proteomics. 22, 100505).