The function of MpWAV3 in Marchantia and its impact on auxin signaling and PIN proteins polarity
SUPERVISOR: Christian LUSCHNIG
PROJECT ASSIGNED TO: Shi YAJING
Unlike animals, plants are sessile organisms and must therefore adapt to their changing environments by modulating their growth, development, and morphology. Central to this adaptability is auxin, a dynamically and asymmetrically distributed plant hormone that plays a pivotal role in numerous growth and developmental processes. The directional transport of auxin, primarily mediated by PIN-FORMED (PIN) efflux carriers, underpins its asymmetric distribution. Specific PIN proteins are polarized to distinct plasma membrane (PM) domains within different tissues, thereby directing the flow of auxin and orchestrating developmental patterns and adaptive responses.
Recent studies have identified WAVY GROWTH3 (WAV3) and its homologs (WAVHs), members of the RING-finger ubiquitin E3 ligase family, as key regulators of PIN polarity and root gravitropism in Arabidopsis thaliana. Furthermore, in rice, the WAV3 ortholog OsGLS1/OsSOR1/OsMHZ2 was suggested to directly control ubiquitination and degradation of OsPIN2. Beyond regulating PIN polarity, WAV3 also mediates the turnover of non-canonical Aux/IAA proteins, highlighting an additional, distinct role in auxin signaling. These findings suggest functional diversification of WAV3 proteins during plant evolution.
In the early-diverging land plant Marchantia polymorpha, only one canonical PIN protein (MpPIN1/MpPINZ), one non-canonical Aux/IAA (MpNCIAA), and a single WAV3 family member are present. This raises key questions: What is the function of MpWAV3? Does it influence MpPIN1 polarity and/or MpNCIAA turnover? Has MpWAV3 undergone functional innovation since the early evolution of land plants?
To address these questions, this study will explore the role of MpWAV3 in regulating PIN polarity and non-canonical Aux/IAA turnover, using a combination of functional genomics, genetic, and bioinformatic approaches. Through this investigation, we aim to uncover how functional innovation within the WAV3 family may have contributed to plant morphological evolution and enhanced adaptability to diverse environmental conditions.
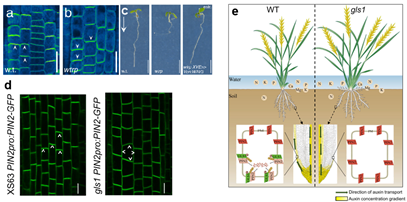
Figure 1. a and b: PIN polarity in wild type (w.t.) and wav triple root meristems; c: Phenotype of wild type (w.t.), wav triple (wtrp) and wav triple XVE»Ven:WAV3. Agravitropic root growth of wav triple is rescued by induced expression of Ven:WAV3; d: Localization of PIN2-GFP in the primary root tip from 7-day-old seedlings of PIN2pro:PIN2-GFP in XS63 and gls1; e: A working model for the regulation of root system architecture in rice by OsGLS-OsPIN2.
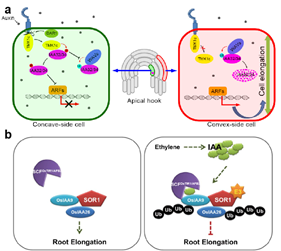
Figure 2. a: AtWAV3 interacts with the non-classical Aux/IAA proteins AtIAA32/34, targets them for ubiquitination and degradation, thereby influencing the formation of the apical hook; b: OsSOR1 is a potential regulator that modulates auxin-mediated ethylene response at the auxin signaling level by interacting with the non-classical Aux/IAA protein OsIAA26, thereby inhibiting root elongation.